
Para poder comprender la escencia de esta nueva propuesta anatómica-funcional de las vias del control del movimiento, primero resumiremos las vías canónicas.
VÍAS CANÓNICAS
- Durante casi cuatro décadas, el modelo clínico predominante sobre los ganglios basales se ha basado en la existencia de dos vías principales y opuestas: la vía directa (Go), que facilita el movimiento, y la vía indirecta (No-Go), que lo inhibe.
- El equilibrio entre ambas sería esencial para el control motor normal, mientras que su desbalance daría lugar a trastornos hipocinéticos, como la enfermedad de Parkinson, o hipercinéticos, como la enfermedad de Huntington.
- Las entradas corticales a las vías canónicas se originan en gran medida en áreas sensorimotora y de asociación del neocórtex, y van hacia zonas dentro de la matriz del estriado.
VÍA DIRECTA (GO)
- Esta vía involucra neuronas de proyección estriatal (dSPN) que expresan el receptor de dopamina D1 en el estriado.
- Se proyecta directamente al globo pálido interno (GPi) y a la sustancia negra pars reticulata (SNpR), continuando en última instancia hacia los objetivos de salida talamocortical y del tronco cerebral.
- Generalmente se considera que esta vía promueve el movimiento.
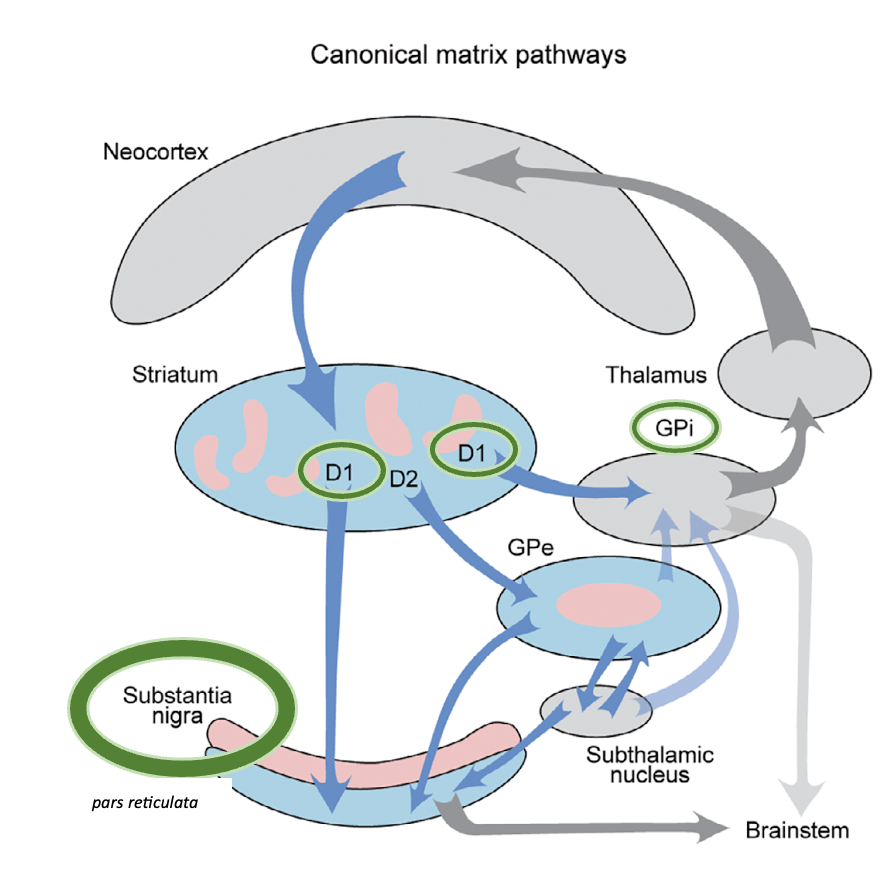
VÍA INDIRECTA (NO GO)
- Esta vía involucra neuronas de proyección estriatal (iSPN) que expresan el receptor de dopamina D2 (iSPN) en el estriado.
- Se proyecta al globo pálido externo (GPe), que luego se proyecta hacia el núcleo subtalámico. El núcleo subtalámico, a su vez, se proyecta hacia el GPi y SNpR, y el GPi luego se proyecta hacia objetivos talamocorticales y del tronco cerebral.
- Generalmente se considera que esta vía inhibe el movimiento.
- El diagrama señala que la vía canónica D2 evita en gran medida la ‘zona central’ rosa del GPe, que es un objetivo para las vías estriosomales (que explico más adelante).
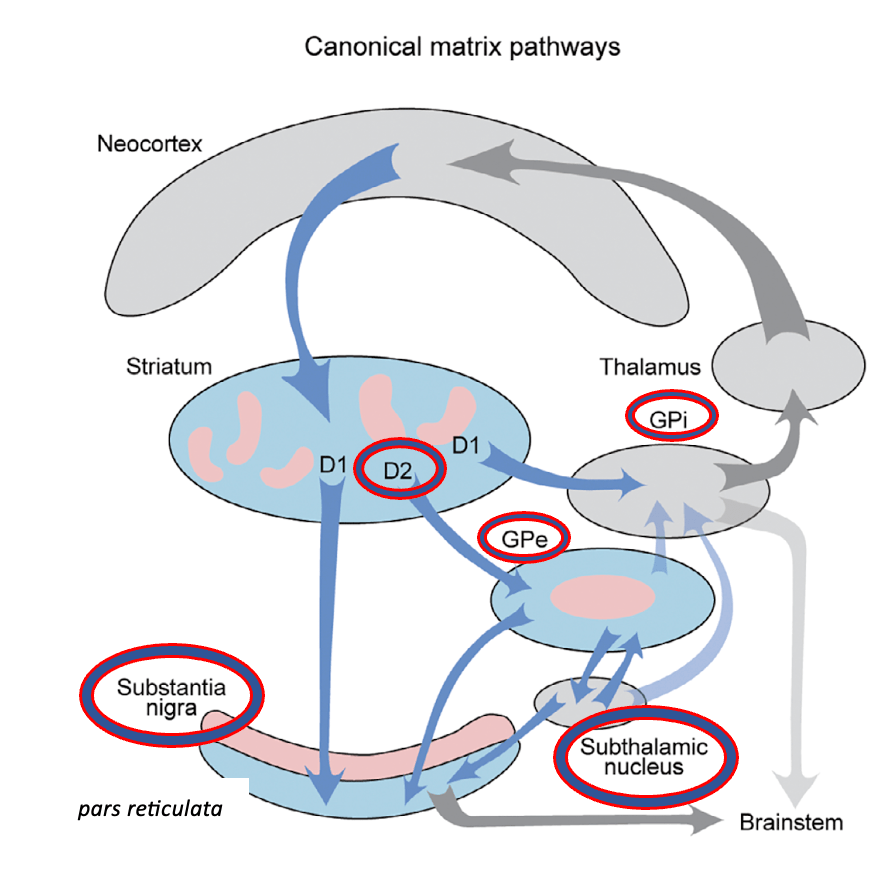
En este artículo, sin embargo, se explica que estudios recientes han desafiado esta visión simplista.
Para ello, lo primero a comprender es que el Globo pálido externo (Gpe), más que un simple intermediario en el control del movimiento en los núcleos de la base (relevo de la vía indirecta), tiene dentro de sí un complejo circuito local y de entrada-salida, y parece que actúa como un nodo autónomo y crítico en la regulación del comportamiento motor y motivacional.
Asimimo, se describe un sistema paralelo de vías “no canónicas”, originadas en los estriosomas (un compartimento del estriado que representa cerca del 20% de su volumen), este sistema es el sistema estriosomal.
Estas vías reproducen la arquitectura directa e indirecta, pero con un destino distinto: las neuronas dopaminérgicas de la sustancia negra pars compacta.
A diferencia de las vías canónicas, las entradas corticales al sistema estriosomal (mostradas por flechas rojizas) provienen principalmente de regiones del sistema límbico, como la corteza cingulata anterior pregeniculada y la corteza orbitofrontal caudal. Estas regiones están implicadas en la toma de decisiones bajo conflictos y procesos motivacionales .
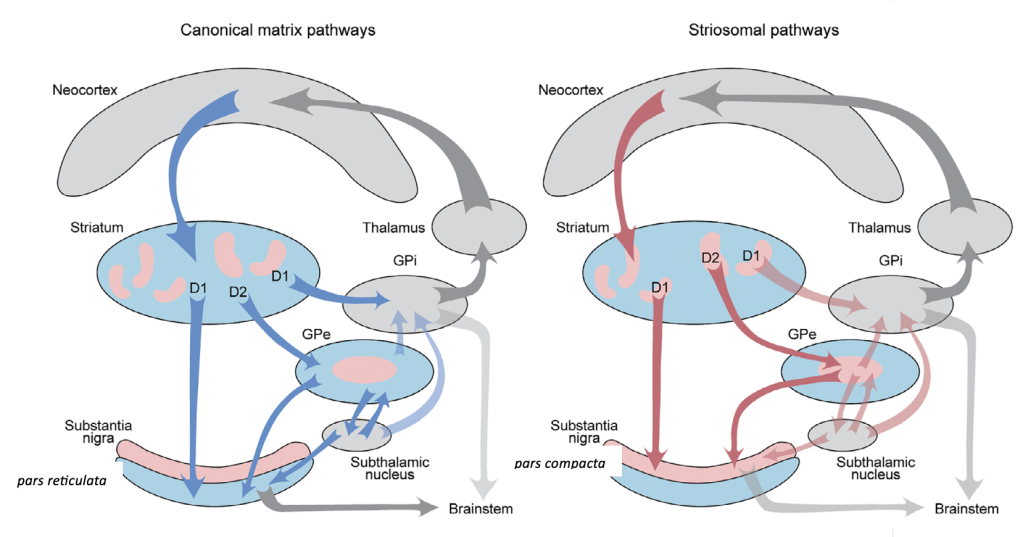
VÍA ESTRIOSOMAL DIRECTA
La vía estriosomal directa (S-D1) se origina en las neuronas espinosas estriosomales que expresan receptores D1.
- A diferencia de la vía directa clásica (matricial), que promueve el movimiento proyectándose al GPi y SNr, la vía S-D1 establece una conexión monosináptica directa con las neuronas dopaminérgicas de la sustancia negra pars compacta (SNpc).
Su estimulación tiene efectos opuestos a la vía directa canónica:
- Disminuye el movimiento.
- Reduce la liberación de dopamina en el estriado.
Esto sugiere que la vía S-D1 actúa como un mecanismo de modulación homeostática, capaz de frenar la actividad dopaminérgica cuando la vía directa clásica está sobreactivada, aportando un control fino sobre la motivación y el comportamiento
VIA ESTRIOSOMAL INDIRECTA
La vía estriosomal indirecta (S-D2) se origina en las neuronas espinosas estriosomales con receptores D2.
En lugar de seguir el trayecto clásico de la vía indirecta, estas neuronas se proyectan hacia una zona central especial del globo pálido externo (GPe), diferente a la que recibe las proyecciones de la vía indirecta canónica.
Lo sorprendente es que su función es inversa a la vía indirecta clásica:
- Aumenta el movimiento, en lugar de inhibirlo.
- Puede contribuir a la liberación de dopamina al modular indirectamente la actividad de la SNpc.
En conjunto, la S-D2 parece actuar como un mecanismo compensatorio o energizante, capaz de contrarrestar un exceso de inhibición producido por la vía indirecta canónica, y vinculándose además con procesos de motivación y toma de decisiones bajo conflicto.
Este hallazgo redefine nuestra comprensión de los núcleos basales, otorgándoles un papel no solo motor, sino también en la motivación, la toma de decisiones bajo conflicto y los síntomas no motores de enfermedades neurodegenerativas.
En la práctica, abre la puerta a terapias dirigidas de manera más precisa: desde intervenciones celulares específicas en el GPe hasta estrategias que modulen selectivamente estas vías estriosomales para restaurar el equilibrio motor y emocional en pacientes con Parkinson o Huntington.
En conjunto, estas “sorpresas” nos invitan a mirar los núcleos basales no solo como un circuito de marcha y parada, sino como un sistema flexible y homeostático, clave para integrar movimiento, aprendizaje y emoción.
BIBLIOGRAFÍA:
Graybiel, A. M. (2025). Surprises from the basal ganglia: Stop and go have new meaning. Movement Disorders. Advance online publication. https://doi.org/10.1002/mds.70008

Leave a comment